| Back to 2nd page (with synopsis) |
p.
03 of 11
|
|||
|
|
||||
| Back to last page |
Next
page
|
|
||
The parent rock has already been described (cf. Darkot & Erinc 1961, Planhol 1956b, Poisson 1977). Pedological investigations in Turkey, including the area under investigation, have been carried out by Oakes & Arikök (1954, after Wilbrandt 1974). For the particular problems relating to agricultural soils in Turkey, reference is made to the com-prehensive study of Christiansen-Weniger (1970). According to Oakes (1954: 57) the soils of the sinter terrace belong to the 'red prairie soils'. Following the morphological classification system of Schroeder (1978) and the description of the above-named soil type in North and Middle America (Gansen 1972: 224/225), for the specific area under investigation preference is given to the soil-type classification of Giesecke (1930, after Christiansen-Weniger 1970: 103). The whole of the southern coastal strip is regarded as Mediterranean skeletal soils with Terra Rossa. These are climate-phytomorphic soils (Schroeder 1978: 105) of variable humidity of the sub-tropics, which frequently occur in the Mediterranean area as relict soils of Pleistocene variable climates (Klinge 1958). Recent Terra Rossa formations occurring on flat, well-drained limestone sub-soils under extreme soil climates (heating, cooling, wetting and desiccation) could also be observed in the area under investigation. The destruction of protective vegetation by extensive agricultural activity leads to extreme soil climates and also to the loss of red soil, which is particularly susceptible to erosion.
Investigations of soil profiles and soil horizons were not carried out in connection with the phytosociological studies. In this context the work of Zech & Cepel (1972) is mentioned, who have investigated the 'Relationship between soil and relief properties and the growth of Pinus brutia in S-Anatolia'. The test areas with the nos. 39 to 42 (Antalya-Kepezalti) of Zech & Cepel are located in a Pinus brutia-forest only a few kilometres away from Nebiler. In view of the homogeneity of the pedogenesis over travertine (Zech & Cepel 1972) the data can be regarded as representative of the study area outside the settlement, even though the humus layer and the A/B-horizon in the area of the Macchie are less deep. Thus the humus layer is 2 to 7cm in depth under Pinus brutia, or the individual bush complex, but seldom more than 1cm in clearings. The A/B-horizon is often broken by the parent rock, reaching a depth of 0 to 30cm with a skeletal proportion of 30-50vol%. Although, as already mentioned, travertine is a homogeneous parent rock with deep clefts and fissures filled with red soil, rooting to a depth of 120cm is possible (lufro 1975: 14). The soil profile descriptions of Zech & Cepel show an increase in hydrogenion concentrations with depth, as already described for Mediterranean red soils by Braun-Blanquet (1928, from Lundegardh 1949: 65). With an average pH-value of 6.65 in the A-horizon (0-10cm) and 5.7 in the B-horizon (10-30cm), the shallow Terra Rossa soils show a weak to moderate acidic reaction.
The relatively high humus layer of 2 to 7cm under even sparse shrub complexes in the Macchie with unfavourable site conditions is understandable in view of the long periods of desiccation of the upper soil layers. Although the pH-values of the humus A-horizon are favourable for micro-organisms, the decomposition of litter is not controlled primarily by the acidity of the upper soil layers, as it is in the humid regions of central Europe, but by the length of the dry season (Zech & Cepel 1972: 36).
Such a sequence of floral composition and typical formations could also be related to certain site factors in the Mediterranean area (cf. Oberdorfer 1965, Knapp 1971, Raus 1979a,b, Naveh & Whittaker 1979). The localised formations are generally the products of development series such as occur under natural conditions, but are in the vicinity of settle-ments due to a variety of activities.
In order to describe the dynamics of vegetation and the floral composition in the vicinity of this small Turkish settlement, and to allow phytosociological comparison with other results, primarily from the Aegean area, use is made of the dimensions proposed by Sukopp (1968, 1969) of intensity, extent and duration of influence. However, for at least two reasons this is only possible with some reservations in the area under investigation. Firstly, little attention is paid by plant sociologists to ecologically relevant site factors, and in particular to their variation over time, a fact pointed out by Walter & Breckle (1983: 125ff). Secondly, the causes of vegetation development in one area are frequently not transferable to another. Due to the scant information available about the history of the village, and conflicting statements about the current economic situation, official information from Antalya was compared with information from villagers and approximate values were determined.
The extent and intensity of human influence on vegetation doubtlessly varies in accordance with the socio-economic system and its environment. For example, in view of their extensive irrigation systems, a larger population can be assumed for the Greek-Roman period on the lower travertine terrace than in the early Middle Ages (Kündig-Steiner 1974). There has only really been an intensification of anthropozoogenic influence with the industrialisation of Antalya over the last 100 years and the development of the infrastructure on the Pamphylian plain, together with the rise in the numbers of sheep and especially of goats.
Although the human influence on the landscape cannot be quantified historically, it is not unreasonable to assume that the traditional social structure has been relatively continuous, since these have changed appreciably only in recent years in other, similar settlements.
The scattered structure of the village, and information from the villagers and the staff of the survey office of the province capital Antalya, give reason to suppose that the village is more than 300 years old. The present population numbers some 600 and is rising according to the mayor (muhtar), with only 279 entitled to vote, and thus older than 21 in 1980. Although there are no statistical records, the existing stock of old buildings, and the relatively small number of new buildings support the opinion expressed by villagers that the population has not suffered any wide variations. The village of Nebiler has some 25 small farmers' families whose livestock and land ensures an adequate income throughout the year.
The figures provided for land and livestock vary widely, and it is certain that villagers have little interest in clarification, at least as far as areas under cultivation are concerned. There has been no land registration, due among other things to the fact that views are divided on the exact distinction between forest and agricultural lands.
As a rule even small families have a garden, with some goats and sheep. Livestock of the village was said to comprise approximately 1.500 goats, 3.000 sheep, 150 cattle and 50 horses, mules and donkeys. There is an observable trend away from sheep and goats towards cattle.
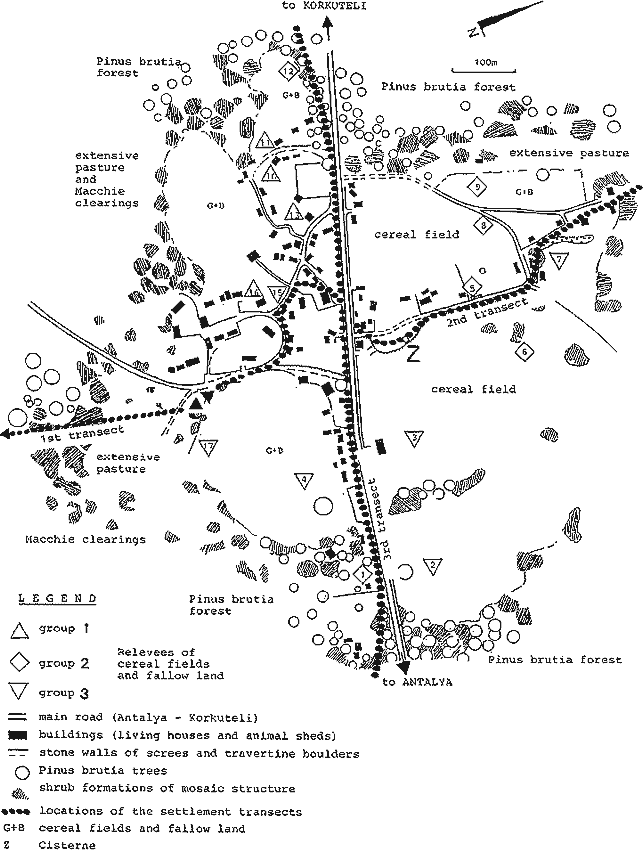
In Nebiler dry agriculture is practised on uneven land in close proximity to the village. Fields often protrude into the Macchie (see Fig. 4). Animal husbandry provides the main source of income for the small farmers, in addition to which wheat and olives are produced solely for personal consumption.
Sowing is in November and the harvest follows in May. In addition to the few olive trees, there occasionally grow carobs (Ceratonia siliqua), figs (Ficus carica) and almonds (Prunus amygdalus). Dung is not used to fertilize the gardens or fields, but is sold to banana plantations or the market gardeners of Antalya.
It should be noted that during field work in summer 1979 near Nebiler, other herds of goats were encountered in addition to those belonging to the village.
Current use of the Macchie and the local forest is primarily for foraging, and only secondarily as a source of fuel. Firewood is obtained mainly by coppicing the strawberry tree (Arbutus andrachne) and from the mock privet (Phillyrea latifolia) as well as from old pine trees (Pinus brutia) some distance from the village. The situation has been eased considerably by recent guaranteed state deliveries of firewood from forestries.
It was not possible to establish whether the areas around Nebiler were also grazed on a regular basis by passing nomads, as mentioned by Jahn (1970) for the western upper terrace. Villagers said this was not the case. Nomads (Yuerueks) are said to have passed on the southern edge of the upper terrace until 1965, which is said not to have had adverse effects on the village's grazing. The absence of Yuerueks in recent years can be attributed to changes in animal husbandry. To meet the requirements of the Antalya market, Daglic sheep gained favour over goats (Jahn 1970: 36).
Asked about grazing behaviour, the villagers emphasised the difference between goats on the one hand and sheep and cattle on the other. In addition to grazing the herb layer, goats ate the spring shoots of almost all shrubs of the Macchie, including Calicotome villosa and Sarcopoterium spinosum. Exceptions are the toxic Daphne sericea, D. gnidium and Myrtus communis, and Cistus species. Sheep and cows, by contrast, restrict themselves to the herb layer, and thus to the range-type of vegetation (extensive pasture) and between the shrub complexes (Macchie clearings). From personal observation, however, it seems that sheep are not averse to the young shoots of Phillyrea latifolia, Rhamnus oleoides and even Quercus coccifera.
The radius of action of the grazers varies widely. The cattle, as well as asses and mules, are kept in the immediate vicinity of the village and tended by the children. Preferred areas are to the south between the Macchie and the village perimeter, and to a lesser extent the area to the north.
The vegetation to the west and south of the village on both sides of the high road consists of more or less continuous Pinus brutia forest, with sporadic to dense Macchie undergrowth to the east of the village perimeter. The picture to the north and south offers a sharp contrast, with no pines near the village. Typical vegetation of extensive pastures gives way to a highly sub-divided, Phrygana-like devastated Macchie vegetation, which gives way to Pinus brutia forest far outside the village.
The term Phrygana is used to characterise devastated, often cushion-shaped vegetation with character species of Macchie formations. 'True' Phrygana according to the definitions of Rauh (1975) or Quezel (1981) were not found in the research area.
Ruderal and segetal flora are concentrated in the village itself, and occur with gradually decreasing frequency in the extensive pastures. The depiction of these transitions of vegetation formations dependent on land-use, with their interactions and dynamics is intended to assist an inductive understanding of stages of degradation of evergreen sclerophyllous vegetation, while also providing a basis for comments on aspects of human influence.
In terms of method, the distribution of species and the alteration of vegetation as a result of specific environmental influences can best be registered using the gradient analysis first applied by the Russian geo-botanist Ramensky (1926, 1930, 1932, cf. Sobolev & Utekhin 1982) and further developed by the American ecologist Whittaker (1967, 1973, 1982). This is based on the view that the vegetation of a landscape represents a complex and extended continuous population pattern which corresponds to the pattern of site gradients (Whittaker 1970: 56).
Plant societies, floristically classified by the Zurich-Montpellier school (Braun-Blanquet 1964, Tüxen 1956) in terms of characteristic and differential species, are seen by the Anglo-American school as overlapping integral units of vegetation perceived as a continuum (Goodall 1963). This continuum can, however, be disrupted by anthropogenic disturbances (Whittaker 1970), or can display boundary zones which correspond closely to the borders of the natural environment (Leeuwen 1965, 1966, 1970, Maarel 1976).
The direct gradient analysis requires a definable ecological parameter or a gradually changing ecological gradient. In our case it is assumed that there is a radial decrease in intensity of land-use from the centre of the settlement to the 'Phrygana' and the Macchie (intensity gradient). While the degree of land-use cannot be measured directly, the distance from the notional centre of the village to the first sampling site some 1.200m from the village can be given and correlated with the individual sampling sites and their results. The sampling of the vegetation stands followed Ellenberg (1956), Braun-Blanquet (1964) and Knapp (1971).
Five layers were distinguished along the three transects (cf. Figs. 4, 5 and 6). The tree layer (I = > 3 m) was formed largely of Pinus brutia, the only species able to dominate clearly over the bush layer (II = 1-3 m, III = 0.5-1 m). The Macchie was subdivided to show the group of nano-phanerophytes, as well as areas in which potential macro-phanerophytes remain low. In layer III species already occur which are typical for the Phrygana, or are transitional to this formation. The herb layer is also subdivided (IV = 0.2- 0.5 m; V = 0-0.2 m). The layer up to 20 cm is clearly dominated by thero- and hemi-cryptophytes of the extensive pastures or Macchie clearings, being also spatially distinct from the dwarf shrubs and ecotonic complexes of the Cistus-Micromeria associations of the Phrygana, consisting largely of chamaephytes (cf. Oberdorfer 1954a, Horvat et al. 1974 and Raus 1979a,b).
The dominance and the sociability were estimated in seven and five classes respectively after Braun-Blanquet (1964). The overlapping of the layers meant that the total sum of coverage was higher than the actual total coverage, which gives a good idea of the density of the vegetation.
The postulate of homogeneity for the classification in accordance with Braun-Blanquet (1964) cannot be realised completely in the gradient analysis, since the areas to be surveyed are to be evenly spaced along the environmental gradient. If the variation of populations and associations is to be registered in full, then the sampling areas cannot be placed preferentially in representative plant associations, since the method of exclusively homogeneous sampling areas is inappropriate for answering syndynamic questions (cf. Goodall 1963, 1973, Barkman 1968, Spatz 1975, Walter & Breckle 1983).
In order to register the entire complex of the Macchie, with residual Pinus brutia stands, the adjacent stages of degradation and its transition to ruderal and segetal vegetation, areas of 200m² (20 x 10 m) generally 15m apart from each other were examined. The 1st transect (cf. Figs. 4 and 5) was begun at the edge of the upper sinter terrace at 235m above sea level some 1.200m SE of Nebiler, leading north-west to the margin of the settlement.
The choice of an area of 200m² in the devastated Macchie was derived from the species number per area-graph after Braun-Blanquet (1964) (cf. Knapp 1971: 33). The herb layer was determined for two areas of 20 m² within the larger area. Within the settlement area (transects 2 and 3), sampling was necessarily restricted to smaller plots, but their spacing remained much the same.
In order to register the ruderal and segetal flora and their distribution, additional surveys were made outside the transects on and around the harvested arable fields. A valuable addition to the vegetation surveys in the settlement area (cf. Willems 1983) proved to be soil samples, from which information about the current seed bank could be obtained by germination tests, with indications of the seasonal (Oberdorfer 1954b) plant associations.
In the course of the survey a considerable number of plants were collected for determination with the Flora of Turkey and the East Aegean Islands (Davis 1965-1985).
The emphasis on the dynamics of vegetation for the tabular classification of the vegetational mapping makes it necessary to organise the entries in the lists according to their position on the transect and to group the species corresponding to similar population densities. In order to be able to judge beyond the individuality of species in terms of their variation with grazing intensity (cf. Walter & Breckle 1983), the centres of distribution of character species (CaS) group combinations, or in the syntaxonomic sense of associations or their fragments, the taxa found along the transects are assigned to groups of character species.
These groups are determined from a comparison of the phytosociological tables and from various group listings of the authors mentioned below. The tables, which are not intended to be a syntaxonomic classification, should only be understood as a deductive determination of groups with character species (abbreviated in the following text as CaS).
This approach makes it possible to portray stages of vegetation and to compare degrees of relationship of 'associations' directly.
Only sporadic floristic-sociological records of the coastal region of south-west Anatolia were available for comparison, so that for the synsystematic and subsequent syntaxonomic evaluation of the taxa found, in order to identify character species, investigations from else-where in the east Mediterranean had to be drawn on for comparison, in particular the work of Oberdorfer (1952a: North Aegean herb and dwarf shrub vegetation, 1956: Canary Islands), Raus (1979a,b: East Thessalia), Lavrentiades (1969: Rhodes), Dafis & Landolt (1975, 1976: Greece), Knapp (1965: Kephallinia), Horvat et al. (1974: Vegetation of SE-Europe), and for the ruderal and segetal vegetation Lohmeyer (1969, unpubl.), Lohmeyer & Trautmann (1970) and Oberdorfer (1954b).
Even if, as Raus (1979b: 20) says, 'heterotonic' tables which include transitional syn-dynamic states following the initially defined approach are not suited for syntaxonomic evaluations in the sense of Braun-Blanquet (cf. Tüxen 1972, 1974, Quezel 1981), they nevertheless allow delimitation between overlapping associations in those areas where species groups are withdrawing and others are entering (Whittaker 1973: 40, Maarel 1976: 420). The spatial dynamic aspect emphasised by the chosen means of presentation clearly shows the hierarchy of relationships of character species groups in the formation of coenoclines (Noy-Meir & Whittaker 1977) related to land-use gradients.