| Back to 2nd page (with synopsis) |
p.
07 of 11
|
|||
|
|
||||
| Back to last page |
Next
page
|
|
||
A further human influence on the diversity is apparent in the extensive pasture type of vegetation, where the extreme grazing has negative effects on species diversity (cf. Schuster 1979, after Pfadenhauer 1976).
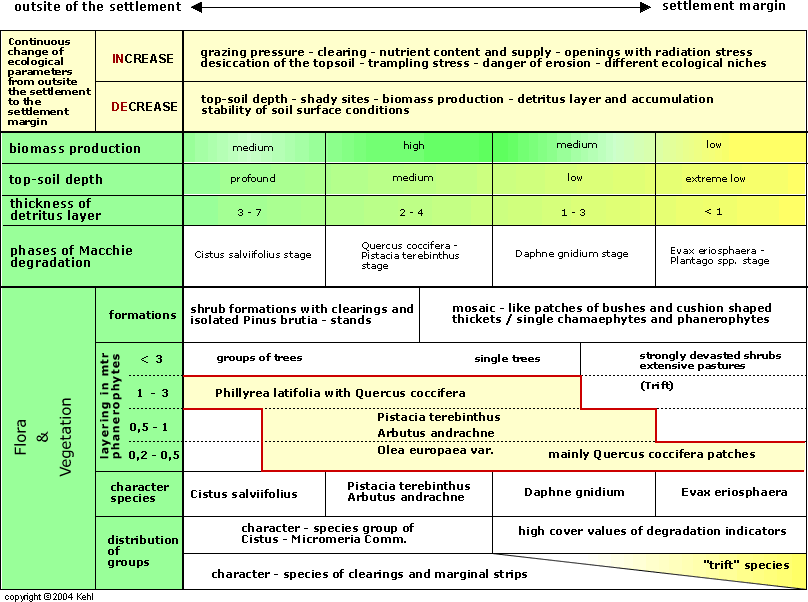
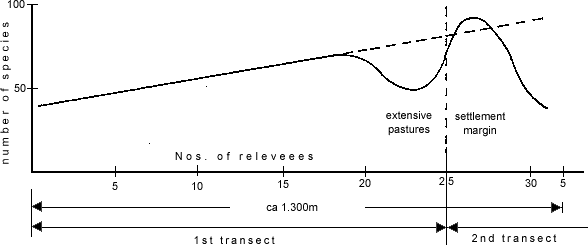
The continuous change of the floristic and structural features of the vegetation, which can be regarded as a degradation sequence, is the result of an extensive, pre-industrial mode of production, which for grazing had the desirable effect of increasing biomass production. The average level of influence which allows a maximum of communities (Schuster 1979: 383) is realized in the region of the devastated Macchie with a high proportion of extensive pasture species, which could theoretically extend right to the edge of the village, as an extrapolation of the diversity graph shows (cf. Fig. 8).
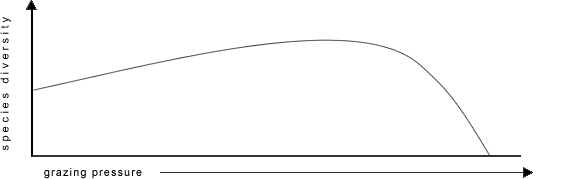
Field experiments on typical Phrygana with Quercus coccifera in Southern France, comparing the dynamics of vegetation and the effects of use, showed a considerable increase in biomass production as a result of moderate use (Poissonet et al, 1978, Fay et al, 1979, Godron et al. 1981). However, the measure of the average use appropriate to achieve the highest possible diversity and biomass production is dependent on the specific vegetation of the production landscape. Thus Whittaker (1979), and Naveh & Whittaker (1979) report the differing effects of grazing pressure on the Mediterranean vegetation in Israel and the USA. An increased level of grazing on Israelian Batha led over a wide range to increased species diversity (cf. Fig. 7), whereas comparable levels in the USA decreased the diversity.
In addition to the reasons noted in the introduction (chapter 1) to the regenerative capacity of Mediterranean vegetation, Naveh (1975: 206), Naveh & Whittaker (1979) and Houerou (1981) see the historical development there of species which are adapted to grazing as a major difference to comparable vegetation types in the USA. An additional factor, especially in the eastern Mediterranean is the heterogeneous topography with considerable differences in altitude, offering a wide range of differing habitats. Many of the species occurring in the Macchie and the extensive pastures are thus almost pre-adapted and form a large reservoir of potential invaders for the new sites created by grazing (cf. Heyn 1971: 191).
The complex pattern of species density demonstrated well on the first transect shows intermediary distribution curves for many species (cf. Fig. 9), which indicates an overlapping and differentiation of ecological niches (cf. Whittaker 1982). The steady succession of emerging and disappearing species with an overall increase in the number of species shows the individuality of the species and the existence of a slowly changing ecological factor, in this case the intensity of land use. With Sobolev & Utekin (1982: 76) and Walter & Breckle (1983: 127) it must further be concluded that the individual species show no sociologic links to one another, but only characterise an ecologically defined site. As Whittaker (1982: 34) comments: 'Species can be assigned to ecological groups by similarity of distribution but the limits of such groupings are essentially arbitrary.'
It is especially important for the evaluation of population patterns that the edaphic and topographic conditions were originally almost homogeneous, and that trampled and shallow-soil sites developed only as a consequence of grazing and wood-cutting. The following section analyses the agricultural use and ecology of the individual sites.
species
The continually decreasing cover values for the Macchie character species with relevee 8 is due firstly to the habitual change of the Macchie elements caused by grazing and secondly to the total disappearance of Phillyrea latifolia, which is still well represented in the middle section. Quercus coccifera only reaches the shrub layer sporadically in isolated bush complexes, and has its highest cover value between nos. 16 and 25 in the upper herb layer (IV), where it frequently only forms an ankle-high cushion. The variation curves of some CaS of the Macchie (cf. Fig. 9) prove their varying ability to counter grazing pressure: Phillyrea latifolia, a class and order CaS of Quercetea (-etalia) ilicis. (Br.-Bl. et al. 1951, Horvat et al. 1974), is represented with a high frequency, with the exception of relevee no. 17, but from the start of the transect to relevee no. 9 shows remarkable growth (cf. Fig. 9). If the vegetation curve of Phillyrea latifolia is compared with the distribution of Daphne gnidium, according to Raus (1979: 37) a good indicator of the degree of use, then it is noticeable that their overlap is considerably between nos. 11 and 17 (cf. Table 1 and Fig. 9).

Returning to the distribution of Phillyrea latifolia it can be assumed that this species only partially intrudes into the degradation stadium named Daphne gnidium-phase by Raus (1979) (cf. Table 3 in Raus 1979: 38/39). It has little ability to regenerate under heavy grazing pressure. If this declines then Phillyrea latifolia, just as Quercus coccifera can form thick shrub cushions (see relevees nos. 3-10). The deciduous Pistacia terebinthus, Olea europaea var. and the sporadically occurring Arbutus andrachne have similar distribution concentrations to Phillyrea latifolia.
It is also noticeable that in this section young Pinus brutia are more common, and are also found in all layers of the relatively continuous Macchie. However, it should be noted that there are always smaller clearings around or near old trees of Pinus brutia where the young plants establish themselves.
Of the CaS of the Macchie the Quercus coccifera has the highest population density along the transect. The species, which only forms taller shrubs in the first two relevees, changes its habitus, but its growth and substance production only decrease slightly at the margin of the settlement. Due to its considerable ability to regenerate (cf. Wraber 1952: 268, Markgraf 1958: 163ff, Knapp 1965: 62, Godron et al. 1981: 326ff, and others) the undemanding Q. coccifera still provides a good source of grazing even in extremely dry summers, when almost all other therophytes have succumbed. It is only gradually displaced by heavy grazing and then only occurs in the extensive pasture area as an anklehigh cushion.
Near the escarpment of the travertine terrace Quercus coccifera has a very different appearance. In the first two relevees the low shrub form is missing; the same applies for Phillyrea. This is because of the different impacts of the two important forms of land use, grazing and wood pollarding, on the Macchie vegetation. As emphasised by Greuter (1975: 126) the population density and physiognomy of various character species of this vegetation type react differently to these forms of use.
Grazing occurs up to the edge of the travertine escarpment, increasing towards the edge of the settlement. In the case of wood extraction by coppicing, however, two directions of extraction can be identified, one going from the village, and the other from the path system of the mixed zone A. Wood is taken from established bush complexes as long poles. The bushes regenerate by pole growth, remaining as individual plants growing up to 2-3 m. This applies to Quercus cocdfera, Phillyrea latifolia, Pistada terebinthus and Olea europea var., of which the latter three have their highest population density in relevee no. 8.
The coppicing and grazing from the mixed zone A is relatively weaker than from the settlement margin, as can be concluded from the absence of bitten Quercus coccifera bushes in the relevees nos. 1 and 2, as well as the presence in relevee no. 3 in the lower shrub layer of Phillyrea latifolia. In contrast to Quercus coccifera, Phillyrea latifolia reacts more sensitively to increased grazing pressure and disappears from the lower shrub and upper herb layer.
Pistacia terebinthus and in particular Arbutus andrachne, which are only sporadically found with pole shoots, suffer particular destruction from grazing and, in the latter case also from firewood collection.
If this CaS group is examined only in economic terms, an optimum biomass production in relation to intensity of land-use, then this optimum occurs between the relevees nos. 5 and 10. Pinus brutia occurs only infrequently, and in its shade there are on the whole few species with high biomass production. The diversity on the above mentioned section of the transect, however, lies some 20% above the first relevee. Therophytes make up the largest proportion, but contribute only to a few percent to the biomass production. The phanerophytes, which are more able to withstand competition, but whose ability to regen-erate in this section is countered by the intensity of land use, would quickly close all clear-ings if anthropo-zoogenous influence were to cease. This would be accompanied by a decline in biomass production (cf. the experiments of Poissonet et al. 1978, Fay et al. 1979 and Godron et al. 1981). The regeneration and biomass production of CaS of the Macchie is then decisively influenced by the intensity of the intervention, which was pointed out at an early date by Adamovic (1929: 33).
In the interspecific competition, with the absence of anthropo-zoogenous influence, the much larger numbers of smaller species would lose out to the few CaS of the Macchie with very high biomass production (cf. also Bornkamm 1961 and Walter & Breckle 1983).
degradation and the character species of the extensive pastures and Macchie clearings
In the Oleo-Ceratonion zone of the North Aegean, Oberdorfer (1954a) has grouped all Phrygana formations of the Quercus coccifera area in the class and order Cisto-Micromerietea (-etalia). The comparison of tables by Oberdorfer (1954a), Horvat et al. (1974) and Raus (1979a,b) with the findings at Nebiler showed that the first transect contained a large pro-portion of the order and class CaS of these communities.
Micromeria myrtifolia, a vicarious species of M. juliana, has its focus of distribution in the south-eastern Mediterranean area of Asia Minor (Rechinger 1950: 83) and has a high presence in almost all relevees in the Macchie. Together with Cistus creticus, Fumana tymifolia var., Teucrium divaricatum, Teucrium polium, Thymus revolutus and others, it settles clearings or the borders of shrub complexes.
Although the CaS of the Phrygana only have low abundances and lower population densities in the total species spectrum, they are a good indicator for the degradation stage of the vegetation in the area under investigation (cf. Fig. 5 and Table 8).
Together with the cropped, frequently cushion-like Quercus coccifera between relevees nos. 15 and 20, there are already signs of a devastated Macchie, as is described by Knapp (1965) for Kephallinia in Greece. However, important components of the Phrygana he describes are either absent in the area under investigation, or are only sparsely represented. Sarcopoterium spinosum and Calicotome villosa, both well protected against grazing and important elements of the true Phrygana (Rauh 1975, Quezel 1981), are only marginally involved in the structure of the vegetation. This section on the transect is emphasised because of the habitual similarity of the above-mentioned highly devastated Macchie to the Phrygana-like formation (cf. Fig. 5).
Only minor differences can be observed in the distribution of the CaS of the Phrygana, with one exception. Whereas Fumana thymifolia var. and Teucrium polium are still encoun-tered frequently in the zone of extensive pastures, the occurrence of Cistus salviifolius is restricted to the mixed zone A at the beginning of the transect.
This species, described by Rübel (1930: 98-100) as undemanding and mainly occurring in the east-Mediterranean area, is characteristic on Crete of the transition from the Macchie to the Phrygana. In this context it should be noted that Oberdorfer (1954: 93) sees Cistus salviifolius as a character species of the Cistion orientale alliance (Oberdorfer 1954), sociologically closely related to the Phrygana community. In contrast to the Cistus-Micromeria communities, which settles mainly on degraded red earth soils, Cistion orientale prefers deeper soils with a higher humus content in the upper strata. In terms of development history, Cistion orientale is closely related to the bush communities of Quercion ilicis.
The above mentioned observations of Oberdorfer are in good agreement with my own findings. In mixed zone A the soils are less burdened, deeper, and have a comparatively marked accumulation of debris, particularly around the cleared shrub formations, as was found by Zech & Cepel (1972) in the vicinity of Pinus brutia stands (cf. 2.3). It is noticeable that in the zone occupied by Cistus salviifolius, Micromeria myrtifolia occurs only sporadically, which can be seen as an indicator that the mixed zone A is most distant from the Phrygana in the degradation series.
Near the area of the extensive pastures at the edge of the settlement the main group of the CaS of the Phrygana between relevees nos. 14 and 19 gradually drops out. This transition is characterised on the one hand by the higher population density of the Daphne gnidium already mentioned, and on the other hand by the increased occurrence of the indicators for extreme degradation. The penultimate stage of Macchie destruction is reached directly at the edge of the settlement with the highest population density and cover on eroded, stony and very shallow soil of the extensive pastures.
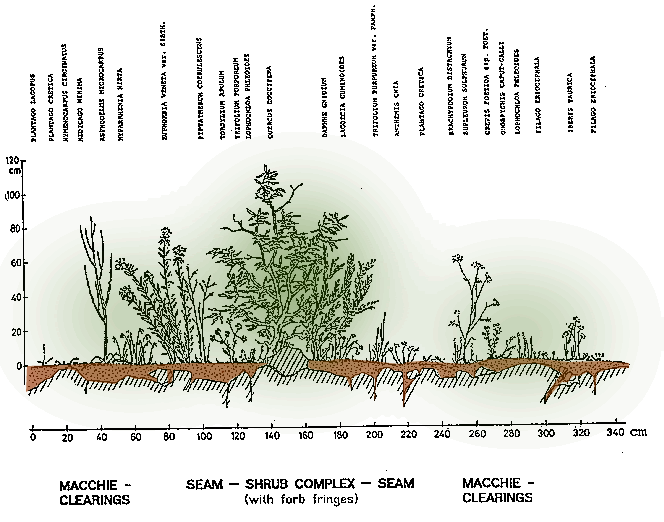
In the shallow-soil pasture zone there is a drop in species diversity, accompanied by a jump in the cover values for a few species. Due to the extreme grazing pressure and the associated soil erosion, conditions have been altered decisively, favouring some dwarf therophytes. The ground was covered with a thick carpet of Evax eriosphaera, only growing a few centimetres high and already withered at the time of the investigation, together with Asteriscus aquaticus, which had also faded. Plantago cretica and Pl. lagopus also had high cover values.
A few ruderal and segetal species, which intrude on the extensive pastures from the inner settlement area, and others which are frequently encountered on nutrient-rich sites or along paths, such as Centaurea solstitialis (cf. Horvat et al. 1974: 128) suggest that the soil on the pastures, though shallow, has in fact a high level of nutrients. This would not be sur-prising, since cows and donkeys frequently graze there.
In the seed bank the geophyte Gagea bohemica was also frequently encountered between nos. 20 and 25 (cf. Table 7), as a geo-element of the steppes (Horvat et al. 1974: 68), as well as Sagina apetala as CaS of Thero-Brachypodietea (Oberdorfer 1954a: 90) and the steppes element Erophila verna (Horvat ibid.: 327).
Therophyte stands, which in the east Mediterranean area often mediate to the Cistus-Micromeria communities at the margins of settlements, are combined by Oberdorfer (1954a: 89) in the alliance Romulion because of their species combination. However, the assignation of the extensive pasture area investigated to this alliance is uncertain, since the important genus Romulea is completely absent. Whether this is an eastern form with Gagea bohemica and Evax eriosphaera, or rather a Poa bulbosa-Plantago lagopus community, can only be answered with additional material for comparison.
There can, however, be no doubt that the entire therophyte vegetation of the extensive pastures and the Macchie clearings can be classified as Thero-Brachypodietea (annual calciferous ext. pasture). Whereas in the North Aegean Oberdorfer (ibid.) only found 50% of the characteristic species of the Th.-Br. known from the west Mediterranean in dominances which determine the association, around Nebiler there were only 50% of those species named by Oberdorfer. An important CaS of this class, Brachypodium distachyum is represented in almost all relevees of the transect with high cover values, with the exception of the mixed zone A and the extensive pastures.
The distribution pattern of the therophytes along the transect allows four ecologically-defined groups to be distinguished, which at the same time demonstrate the gradient of land-use.
In addition to the first group (cf. Table 1) which avoids the denser Macchie and has two distribution focal points, a second group can be recognised, whose species regularly settle with few exceptions in the mixed zone A and the extensive pastures, with a high cover value between the settlement and layer step borders. The first two relevees in the mixed zone A are poorer in therophytes. For the species in the first group the higher levels of shade in the denser Macchie are obviously a limiting factor for distribution, as are the layer step border zone, rich in chamaephytes with more humus in the upper soil levels. Almost all clearings of the bush formations and mosaics of shrub complexes are occupied by species of the second group, which with Psilurus incurvus, Anthemis chia and Plantago cretica are also an essential part of the species spectrum of the shallow-soiled extensive pastures.
The heavily grazed and eroded extensive pasture zone is occupied by the third group with Evax eriosphaera, Gagea bohemica (cf. Table 7) and Astericus aquaticus. It is not possible to say whether Gagea bohemica can be regarded as an indicator of the penultimate stage of Macchie destruction in view of the lack of information about its occurrence on the southern transect section. On the other hand Evax eriosphaera, an east Anatolian vicariant (Rechinger 1959: 86) of the western E. pygmaea, occurring on stony pastures and dry places (cf. Polunin 1980: 448), can serve as CaS of the pasture areas exposed to intense land-use. An observation by Braun-Blanquet (1964: 408) is also instructive in this context, namely that the CaS of the Thero-Brachypodion, to which the west Mediterranean Evax pygmaea belongs, have a high resistance against extreme soil temperatures. It can be assumed that on the extensive pasture area the virtual absence of a shading shrub layer means that the highest soil temperatures are reached there.
In the heavily devastated Macchie with open mosaics of shrub complexes, the third group of the extensive pasture species is neighboured by a further species group of clearings and borders. This group accompanies the Daphne gnidium stage with Hyparrhenia hirta, Xeranthemum annuum, Bupleurum sulphureum and Tordylium apulum (Raus 1979: 37). The latter two groups populate the main distribution area of the CaS for extreme degradation and together characterise the last stage of destruction of Macchie around Nebiler. The species groups indicating the Daphne gnidium stage are mainly followed by species occurring in the Macchie, with intermediate distribution patterns of their populations.