| Back to 2nd page (with synopsis) |
p.
08 of 11
|
|||
|
|
||||
| Back to last page |
Next
page
|
|
||
The observations recorded by Schwarz (1936: 324) on the north-west Mediterranean coast of Turkey, and by Oberdorfer (1954a: 89) in the North Aegean, that the usually im-passable bush stands of the Macchie takes on a mosaic pattern near settlements or their lines of access, and that between the individual shrub complexes the open soil is covered with therophytic vegetation, or species of the Phrygana, also applies to the Nebiler area. The formation of a true Phrygana, with stand formations of thorny dwarf shrubs (Rauh 1975, Quezel 1981), or species characterised by seasonal dimorphism (cf. Orshan 1964, Margaris & Vokou 1982), could not be established in the area under investigation. It was, however, possible to establish degeneration stages of Macchie with marked layer formation, in the lower shrub layer of which some typical representatives of the Cistus-Micromeria community have migrated.
The
suspicion expressed by Turrill (1937), Regel (1943), Zohary (1973) and
also Walter (1956), that Macchie at higher levels from which Pinus
brutia was extracted would not be able to regenerate to a forest,
except after bush fires (Walter 1973: 129, 1975: 77), cannot be confirmed
by my observations made around Nebiler at an average of 250 m above
sea level. Even very sparse Pinus brutia stands with a well
developed and thick shrub layer on a weakly developed topsoil show a high
proportion of seedlings and young plants.
syntaxonomic evaluation
Furthermore, it is generally difficult to draw a clear distinction between CaS of Secalinetea and Chenopodietea. In contrast to Oberdorfer (1954b: 381), who found barely any overlap, Zohary (1973: 636ff) notes a broad overlap of the two communities, which could be confirmed in this investigation. Above all on the harvested fields there is close intermeshing of ruderal and segetal species (cf. Table 5). The co-existence of ruderal, segetal, trampling, extensive pasture and Macchie species (cf. Fig. 11) on dry stone walls between fields and along paths was observed by Schwarz (1936: 381) in West Anatolia. A systematic registration of segetal communities seemed to him to be impossible, as did any generalisation, since 'if at one place certain species seem to be mutually exclusive, the same species may cover the ground elsewhere in harmony, and colourful profusion' (Schwarz 1936: 318). Even though it definitely is possible to recognise focal points with clear mass development in the area under investigation and the species cannot be said to have no diagnostic value, in contradiction to Schwarz's view, his comment does illustrate the fluid borders of the so-called weed communities with those of the Quercus coccifera communities in the margins of the settlements.
The last-named species is also classed by Oberdorfer (1954: Table 6) as order CaS of the Chenopodietalia mediterranea Br.-Bl. (31) 36. The species spectrum of group 2 (cf. Table 5) also includes Thymelaea passerina and Ajuga chamaepitys subsp., whose membership to the 'culm crop-weed' communities is also disputed (cf. Braun-Blanquet et al. 1951: 48, Oberdorfer 1954b: Table 1, Horvat et al. 1974: 315). The virtually isolated occurrence of Hypericum triquetrifolium in the outer margin areas of the fields has been remarked on in chapter 5.8. This normally perennial species (Zohary 1966: 223, Davis 1965-85, Polunin 1980: 336) completes its entire vegetation cycle in one vegetation period in the area under investigation, like summer-annual plants. Hypericum triquetrifolium is spread throughout to the Cyrenaica in North Africa (cf. Rikli 1946: 991), settling on open stony sites of degraded Macchie, and is frequently a companion of the Sarcopoterium communities.
In the east Mediterranean this species, also found on harvested fields and fallow lands, must instead be included in the facultative segetal flora as in Zohary (1973: 649ff). This group includes species which have their natural distribution in the 'primary plant communities', but are able to settle cultivated areas during secondary expansion. It also includes Ajuga chamaepitys subsp. chia and Thymelaea passerina, which in the Danube Basin of Vojvodina (cf. Horvat et al. 1974: 315) can be a CaS of Secalinion orientalis assoc. (cf. also Braun-Blanquet et al. 1951: 47). Further species are Scandix pecten-veneris and Filago pyramidata, the latter being a frequent companion of the Cisto-Micromerietalia Oberdorfer 1954 (cf. Raus 1979: 68 and Table 1 of this paper), in which Braun-Blanquet et al. (1951: 48) sees an alliance character species of the Secalinion mediterranea. Zohary (1973: 649), on the other hand, includes Filago pyramidata in the group of the 'Post-Segetals', which forms the initial stage for succession series on abandoned fields together with e.g., Plantago cretica, Trigonella monspeliaca, Delphinium peregrinum, Consolida hellespontica etc. (cf. Table 5).
In summary, the vegetation surveys of the harvested cornfields and fallow in the settlement area of Nebiler produced only a small proportion of really obligatory segetal flora. According to Zohary (1973), only two important CaS of the 'culm crop weeds' (non-irrigated winter and summer crops) are well represented in group 2. Although there are similarities to the Heliotropeto-Chrozophoretum of Oberdorfer (1954) the assignation to the Diplotaxidion seems problematical, at least for the south-eastern Mediterranean. On the one hand the titular Diplotaxis species are absent, as Oberdorfer (ibid.: 385) emphasised, and on the other hand all the named CaS of this community occur mainly on summer stubble fields, shallow fallow land, and some of them even frequently on rocky sites as companions of the Phrygana (Zohary 1966: 223, 1973: 634ff, 1971, Raus 1979: 60, Polunin & Huxley 1976: 178, Polunin 1980: 336, Horvat et al. 1974: 119).
In this context, attention is again drawn to the mass development of Hypericum triquetrifolium. A clear correlation between higher population density and site conditions could not be established. The majority of the relevees of this group are in the outer eastern margin area of the fields, so that it can be assumed that in particular nos. 2, 4, 7 and 17 lie in areas which have not long been used agriculturally, but this criterion no longer applies to the nos. 3 and 15.
Only long-term observations over several years on the same fields could establish whether the population density is due solely to the diaspora supply of differing farming or fertilizing methods, or to edaphic conditions.
The relevees from the more central settlement areas combined in group 1, with a high proportion of CaS of the ruderal community, are essentially intermediate to the road- and fieldside plant communities of the 2nd transect (cf. Table 2). The relevee no. 13 with high cover values of Heliotropium hirsutissimum and the presence of Amaranthus albus, Marrubium vulgare, Hirschfeldia incana, Chrozophora tinctoria and Tribulus terrestris points to the segetal group 3, the additional occurrence of ruderal species also produces certain similarities with Heliotropeto-Chrozophoretum Oberdorfer 1954. Alliance CaS of the Diplotaxidion however are completely absent.
Whereas no. 15 has Hypericum triquetrifolium as dominant species, no. 16, only a few metres away has Scrophularia canina subsp. bicolor which grows directly next to buildings, on extremely shallow, poorly worked soil. This perennial Scrophularia, frequently encountered on rocky sites (cf. Rikli 1946: 540, Polunin 1977: 312, 1980: 540), and which also occurs as a companion of the Picnomon acarna community along paths (Oberdorfer 1954b: Table 20), characterises a situation at the edge of fields where adequate and sufficient working is hardly possible.
The
relevees nos. 10 and 11 lead over to the CaS combination of communities
growing at the foot of walls, on field and path sides. Hirschfeldia
incana, which is particularly well represented here, spreads out into
the adjacent fields, but it has its highest population densities on the
less-frequented edges of fields, paths and around buildings. At the same
time there is an increase in the proportion of species which find their
way from the nearby extensive pastures and Macchie along the pathways
into the centre of the settlement (cf. the vegetation relevees
of the periphery of the central village open space in chapter
4.5).
In the group of ruderal species (cf. Table 6) there were 11 new species in the seedbank, of which only five CaS of the Mediterranean 'intertilling crop weeds' germinated more than three times in the soil samples, none of which according to Oberdorfer (1954b: 395) led the characteristic ruderal communities established by him for the early spring on the Balkan peninsula. Thus neither Sisymbrium orientale nor S. irio could be determined in the seed bank. On the other hand there were many seedlings of Amaranthus albus, found irregularly in the vegetation surveys, and of Sisymbrium officinale, which in particular was already found frequently in the central settlement area. The high population density of Sisymbrium officinale deserves particular attention, since according to Oberdorfer (1954b: 382) in addition to Lactuca serriola as typical CaS of the Euro-Siberian Ruderetea, Sisymbrium officinale already belongs to the rare species of the southern Balkan. In the area under investigation this species clearly marks considerably eutrophied sites in the centre and to the north of the settlements (cf. Table 2) where the concentration of dwellings and sheds gives rise to an accumulation of dung and kitchen waste at waysides and near houses. If one studies the array of species, and the proportion of nitrophile species, important differences become apparent.
Between nos. 51 and 56 a Hirschfeldia incana-Hordeum leporinum-wayside community forms, with a high proportion of accessory pasture and segetal species, but few of the CaS of Chenopodium murale communities restricted to sites rich in nitrogen. At the foot of walls a character species, Arum italicum, is somewhat more frequently represented, and only the relevee no. 54, with the sporadic occurrence of Chenopodium murale, Chenopodium opulifolium and Marrubium vulgare can be linked to nitrogen enrichment indicating Chenopodium communites of the centre of the settlement.

Hirschfeldia incana, well represented over the settlement area as a whole, is completely absent from the actual centre of the settlement (see Table 2). More nitrophilous species occur such as Chenopodium murale, Chenopodium opulifolium (rare), Chenopodium vulvaria, Marrubium vulgare, Brassica napus, Mercurialis annua, Euphorbia helioscopia, Erodium cicutarium, Amaranthus albus, Stellaria media and Lolium rigidum var., which in their combination clearly show in the vegetation of the central settlement area the alliance Chenopodion muralis Br.-Bl. 1936 of less-frequented ruderal places.
From a floristic-ecological point of view the CaS spectrum of the central settlement area are intermediate to the closely related Euro-Siberian alliance Sisymbrion officinalis Tuexen, Lohmeyer and Preising 50 of the warmer East-European, nitrogen-rich ruderal sites. The Euro-Siberian alliance, with a more predominantly summer life-cycle and a higher proportion of short-lived species (Oberdorfer 1954b: 395, Horvat et al. 1974: 410) extends into the margins of the settlement, in contrast to the Chenopodion muralis alliance, which lasts from spring to late-summer.
As Oberdorfer (1954b) observed on the Balkan peninsula, and Lohmeyer (1985, pers. comm.) on Crete, the CaS of the alliance Chenopodion muralis in Nebiler are tied closely to the actual centre of the settlement, though without any precise border to neighbouring CaS groups. For example the well-developed Mercurialis annua community, on 'dry sites, poor in fine soil' (see Oberdorfer 1954: 390, Table 13) spreads far into the Hirschfeldia-Hordeum community of the settlement margins. However, with its CaS Mercuralis annua, Calendula arvensis and Antirrhinum orontium this community settles the base of walls and pathways, with soils rich in fine soil, humus and frequently covered with travertine rocks, but is also found in the shadow of loose stone walls next to the CaS of the Macchie and the extensive pastures. This society of the 'Steinschutthalden' as Oberdorfer (ibid.) called it, is doubtlessly restricted to nitrogen-rich ruderal sites, but already forms a transition to the communities dominated by Hirschfeldia incana.
A CaS group limited solely to the centre of the settlement could not be found, but an area was identified which was characterised by the presence of Chenopodium murale and by the almost total absence of Hirschfeldia incana, and which can be regarded as the real core area of the Chenopodion muralis alliance. Only here is it possible to recognise slight tendencies to the Chenopodion muralis Br.-Bl. (1931) 36, although the most important CaS of this association are sparsely represented. The reason for the weak formation of this association could be that, according to Oberdorfer (1954b: 395), the main development of this ruderal community only occurs in late summer or early autumn. Another factor could be the lower levels of nutrients, and more extreme dryness than for example on the Balkan peninsula. The seed bank investigations do not support the possibility of the seasonal low development of the Chenopodion muralis (cf. also 5.9), since it provides no significant augmentation of the spectrum of CaS. It can therefore be assumed that the relatively low levels of eutrophication as a function of the density and extent of the settlement are the reason why Chenopodion muralis did not develop fully.
The wayside community reaching nearly to the heart of the settlement, with Hirschfeldia incana, Crepis foetida subsp., Hordeum leporinum, Sisymbrium officinalis and occasionally also Malva nicaeensis could be attributed to the Hordeion murini Br.-Bl. 31 alliance, or to Bromo-Hirschfeldion Lohm. 75.
Hirschfeldia incana and Hordeum leporinum, major components of the wayside communities, and determinants of the aspect in spring, had already died back during the survey in June/July 1979 due to the very hot and dry conditions. Only Malva nicaeensis and Echium plantagineum were still flowering.
As already stated, Hirschfeldia incana characterises the nitrogen-rich sites outside settlements, but can also extend along the waysides into the heart of a settlement, where it is replaced by Chenopodium murale. The CaS of the alliance of Hordeion murini, Hordeum leporinum has a much wider distribution, and can frequently be found with a high presence as CaS of the Chenopodion muralis (cf. Oberdorfer 1954b, Horvat et al. 1974, Lohmeyer 1975, Lohmeyer & Trautmann 1970) and beyond the wayside communities near the settlement is also a good character species of the Onopordion illyrici (Chenopodietalia mediterranea) succeeding the Mediterranean therophytes.
In addition to Hirschfeldia incana (which determines the early summer aspect), Malva nicaeensis, Lolium rigidum, Avena barbata, Urospermum picroides and Cynodon dactylon, according to Zohary (ibid.) Hordeum murinum (?) is also represented as a CaS (on the distribution of H. murinum and H. leporinum see also Faruqi 1980). The vicariant of Malva parviflora, M. nicaeensis, mediates in its distribution in the settlement centre with the Chenopodion muralis, where the relevees 36 and 37 show similarities to the Chenopodio-Malvetum parviflorae of Lohmeyer & Trautmann (1970: 221) for La Palma. Although already occurring together with many CaS of the Macchie and extensive pastures at walls and the edges of fields in the 2nd transect, there are signs in the northern section between nos. 52 and 56 of a Hirschfeldia incana-Hordeum leporinum association. The species spectrum recorded, with only a few characteristic nitrophilous species corresponds to the Hirschfeldia incana-Hordeum murinum community described by Oberdorfer (1965) for Teneriffa or the association Hordeetum leporini Br.-Bl. 36 proposed by Braun-Blanquet (1936) for the western Mediterranean.
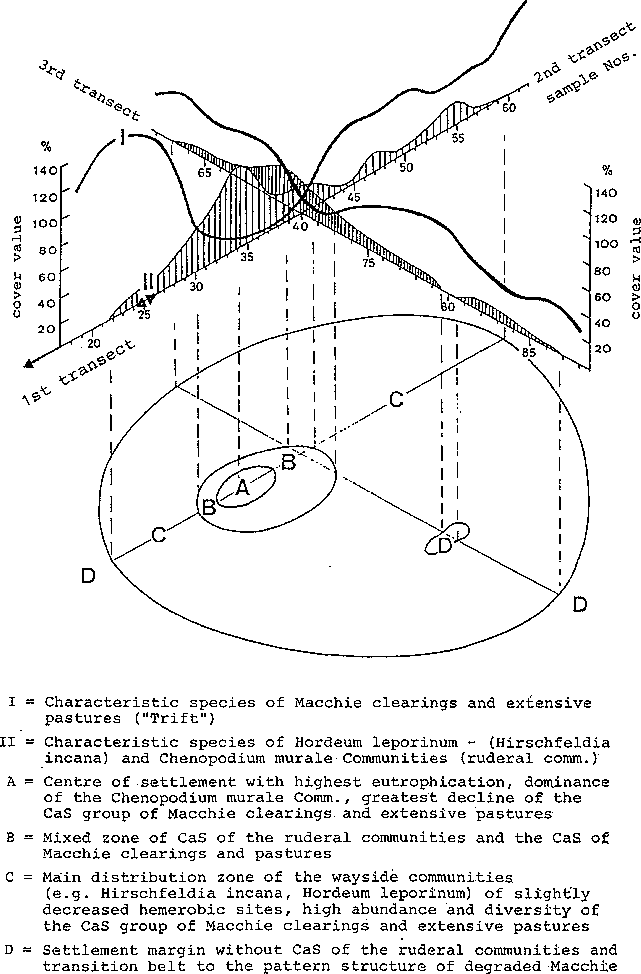
A close interweaving with communities remote from the settlement, or with their CaS groups can be observed on the southern margin of the settlement. CaS of all the groups determined settle on a small area, and only the extensive pasture and trampling species make use of the more compacted and frequented space between wall and path (see Fig. 11).
There is a flowing transition from the trampling community with Cynodon dactylon, Polygonum arenastrum, Matricaria chamomilla, Polycarpon tetraphyllum and Plantago coronopus (see Table 2) to the narrow zone of extensive pasture species and wall foot communities with Hirschfeldia or Chenopodium murale. The CaS of the extensive pastures and Macchie clearings mediate as accessory species between both communities, but can also occupy the entire area between the ways and limiting walls as Poa bulbosa-Plantago lagopus assoc., if the trampling and wall foot communities retreat.
Polygonum arenastrum, an east Mediterranean vicariant of P. aviculare, occurring in close accompaniment with Malva nicaeensis, Mercurialis annua and Euphorbia helioscopia (cf. Table 2, nos. 33-36, and Zohary 1973: 646) characterises nitrogen-rich sites in the settlement, and is! attached to the Chenopodium murale-wayside community. Similar to Polygonum aviculare in Central Europe (cf. Bornkamm & Meyer 1977), Polygonum arenastrum occupies more polyhemerobic than euhemerobic sites in the area under investigation (cf. Sukopp 1972, Blume & Sukopp 1976, Bornkamm 1980). The character species of the extensive pastures are only represented very sporadically outside the central settlement area. Only Cynodon dactylon occurs regularly on waysides over almost all the 2nd and 3rd transect. After Oberdorfer (1954b) it can no longer be used as a characteristic Mediterranean species of the trampling community, since it is widespread around the Mediterranean on dry ground and as a companion of nitrophilous communities as well as therophyte flora (cf. also Lohmeyer 1975: 134, Raus 1979: 54).
Without
Cynodon dactylon and Telephium imperati (see note to Table
2) the CaS group of the trampling communities are closely restricted to
the heavily trodden and eutrophied settlement core. It must also be emphasised
that Polycarpon tetraphyllum after Lohmeyer (1975: 135) does occur
mainly in nitrophilous trampling communities, but that it is frequently
a companion of the stony extensive pastures and therophyte stands (Braun-Blanquet
et al 1951: 218, Oberdorfer 1954: 91) as well as accessory to the segetal
communities (Zohary 1973: 638). The exceptionally high numbers of seedlings
of Polycarpon tetraphyllum (cf. Table 7) in the soil samples
of the extensive pasture area suggests that this species is also widely
spread on the clearings of the Macchie in the spring.